
联系电话
181-7148-1107
181-7148-1107
随着水资源日益匮乏,全球干旱区域不断扩大, 干旱程度日趋严重。 干旱严重影响植物的生长和发 育,如何提高植物的抗旱性已成为遗传育种研究的 重中之重。 目前,关于植物抗旱机制的研究取得了 较大进展,主要包括:活性氧(ROS)的清除、气孔行为、渗透调节、植物激素水 平的调节和光合效率的提高。 干旱可诱导植物产生大量ROS,而过量的ROS积累会对植物产生损害。 植物主要通过自身抗氧化系统清除体内多余的ROS。 植物抗氧化系统包括抗氧化酶系统和抗氧化 剂系统。 抗氧化酶主要有超氧化物歧化酶(SOD)、 过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)、 单脱氢抗坏血酸还原酶(MDHAR)、脱氢抗坏血酸 还原酶(DHAR)和谷胱甘肽还原酶(GR)。 研究抗氧化酶在植物体内的抗旱机制,以及利用基因工程来增强抗氧化酶相关基因的表达,对提高植物抗旱性有着重要的现实意义。
1 干旱胁迫中 ROS 的产生及对植物的损害
ROS 是生物体内活性含氧化合物的总称,主要包括超氧阴离子自由基、过氧化氢、 羟基自由基和过羟基自由基等。 在正常情况下,植物体内的ROS处在一个较低水平,且 ROS的代谢保持动态平衡,不会对植物产生 伤害。 但干旱会明显增加植物体内ROS含量和产 生速率。 例如,随着干旱胁迫时间的延长,金心吊兰和苹果树苗叶片各区域的ROS 含量和膜脂过氧化产物丙二醛(MDA)含量表 现出显著上升的趋势, ROS产生速率也逐步增加,说明干旱胁迫会促进ROS的积累。 ROS大量积累会导致ROS代谢平衡被打破,从而造成植物氧化损伤。 这些损伤主要包括3方面:(1) 对生物 大分子的伤害,如引起蛋白质结构发生改变,导致蛋白质肽链断裂,进而使酶失活;(2)损害细胞的结构 和功能,如抑制叶绿体的正常发育,影响光合作用速率;(3)改变细胞膜通透性,导致 MDA含量迅速增加,改变细胞膜脂质的结构和运动状态,膜脂质过氧化,破坏膜的完整性等, 严重时甚至导致细胞死亡。
2 SOD、CAT 和 APX 在抗旱过程中的功能
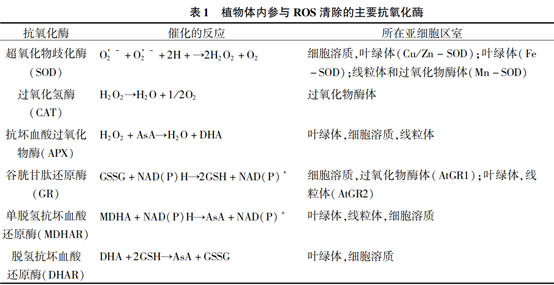
植物抗氧化酶主要种类及其催化的反应、所在亚细胞区室如表1所示。 SOD、CAT 和 APX3种酶广泛存在于植物细胞内,它们通过直接清除ROS以调控ROS水平,并把ROS 转换成活性较低和毒性更小的有害物质。 它们因为直接与ROS反应,所以被看作是细胞内ROS的传感器。
2.1 SOD 在抗旱过程中的功能
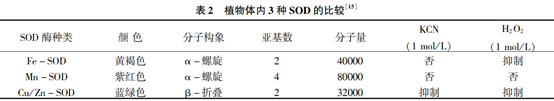
SOD 是一类广泛存在于植物细胞内且容易被ROS诱导的金属酶。 在干旱胁迫下,SOD 作为抗氧化的第一道防线主要通过清除超氧阴离子自由基以减少哈伯-韦斯反应生成更活跃的羟基自由基的风险。 SOD 催化的歧化反应比自发的反应速度要快10000 倍。 SOD 根据金属离子的不同可以分为 3种:Cu/Zn -SOD、Mn -SOD 和 Fe -SOD,它们的生化特征如表 2 所示。 当干旱胁迫诱导 ROS 增加时, SOD 的活性和含量也随之增加。 李萍等对不同品种的蚕豆做了干旱胁迫试验,结果表明:SOD 活 性随着干旱胁迫的增强呈现出先升后降的趋势,且抗旱品种的 SOD 活性增加程度大于敏感品种。
2.2 CAT 在抗旱过程中的功能
CAT又称触酶,为同源四聚体,含有血红素,主 要存在于植物的过氧化物酶体和乙醛酸循环体中。 CAT 可直接分解和催化分解 H2O2 ,但它与 H2O2 的 亲合力相对较弱,所以 CAT 在清除逆境胁迫产生的 H2O2 时有一定局限性。 植物体内的 CAT 可分为 3 类:CAT1、CAT2 和 CAT3。 其中,CAT1 能清除光 呼吸过程产生的 H2O2 ,主要存在于光合作用组织 中;CAT2 受 UV-B、病原物和臭氧诱导,在植物抗 逆中起重要作用,主要存在于微管组织中;CAT3 主 要清除乙醛酸循环中脂肪降解产生的 H2O2 ,在种子内表达丰富。 研究发现,干旱胁迫能够诱导 CAT 基因上调以促进 H2O2 的分解。 对烟草进行干旱处 理时发现,CAT1 基因表达量随着干旱时间的变化而 发生改变:前期轻微增加,中期达到最高峰,是对照 的 20 倍,后期表达量明显下降至与对照相近。 这一 研究结果说明烟草 CAT1 基因参与了植物抗旱信号传导过程。
2.3 APX 在抗旱过程中的功能
APX 主要存在于叶绿体基质中,是一种由卟啉与肽链构成的血红蛋白,是清除叶绿体中抗坏血酸-谷胱甘肽(AsA -GSH)循环所产生的 H2O2 和催 化 AsA 氧化的关键酶。 APX 与 CAT 不同之处在 于 APX 与 H2O2 的亲合力较强,但需要 AsA 作为底 物。 根据 APX 在植物细胞中定位的不同可分为:细 胞质 APX( cAPX)、类囊体 APX ( tAPX)、微体膜 APX (mAPX)和叶绿体基质可溶 APX( sAPK)。 有研究表明:在干旱胁迫下,小麦抗旱品种的 cAPX 活性增加程度明显高于不抗旱品种。 此外,枣树 ZjAPX 基因在干旱胁迫下诱导表达,在一定时间和 浓度范围内,随着干旱程度的加大表达量增加,说明 APX 参与了减少干旱给植物带来的损伤。
3 GR、MDHAR 和 DHAR 在抗旱过程中的功能
另一组酶为 GR、MDHAR 和 DHAR,它们参与 调控抗坏血酸(AsA)和谷胱甘肽(GSH)合成及还原 的过程,从而平衡植物细胞的氧化还原状态。 这 3 种酶催化的反应和所在亚细胞区室见表 1。 在干旱胁迫下,由于 ROS 的大量积累,会引起抗氧化酶 在转录水平和蛋白水平上的上调以达到调控 ROS 水平的目的。
3.1 GR 在抗旱过程中的功能
GR 是一种黄素蛋白氧化还原酶,主要分布于 叶绿体、线粒体和细胞质中。 在干旱胁迫下 GR 的 活性增强,该酶一方面以氧化型谷胱甘肽(GSSG)为 底物催化生成 GSH,有利于 AsA 的再生和激活 CO2 同化中的酶类;另一方面还可以增加 NADP + /NAD- PH,保证了 NADP +的供应,确保 NADP +对来自光合电子传递链电子的充分接收,使超氧阴离子自由基的形成受到抑制。 宋贵方等从抗旱品种陆地棉中克隆得到谷胱甘肽还原酶基因(GhGR),发现在受到干旱胁迫时叶和根中的 GhGR 表达量上调,并且表达量随着胁迫加重而增加,说明GR在陆地棉抗旱信号传导中发挥一定作用。
3.2 MDHAR 和 DHAR 在抗旱过程中的功能
MDHAR 和 DHAR 都是AsA代谢途径的关键酶。 在干旱条件下,MDHAR 和DHAR活性变高,并分别将AsA的氧化产物单脱氢抗坏血酸(MDHA) 和脱氢抗坏血酸(DHA)还原再生成AsA,对维持植物细胞内AsA的平衡和氧化还原平衡起着重要作用。 有研究表明,干旱胁迫激发了冰草叶片中抗坏血酸防御系统,使清除H2O2的 APX和加快AsA循环再生的MDHAR和DHAR酶活性上升,从而维持叶片抗坏血酸水平和缓解干旱对植物的伤害。
4 研究展望
植物抗氧化酶系统能够有效清除细胞内 ROS, 是植物抗旱保护机制的一个重要组成部分。 目前, 关于抗氧化酶在植物抗旱信号传导中的作用机制的 研究取得显著进展,但仍有许多尚未解决的问题。 主要问题和具体对策如下:(1)目前克隆出的抗氧 化酶相关抗旱基因还不多,需要挖掘更多抗氧化酶 基因资源以用于植物抗旱基因工程;(2)目前植物 抗氧化酶抗旱基因工程多为单基因转化,而植物清 除 ROS 是多种抗氧化酶协同作用的结果,单一提高 某个抗氧化酶基因的表达,有一定效果,但并不明 显,甚至会产生反作用,因此多基因转化能更显著地 提高植物的抗旱性;(3)目前对抗氧化酶转基因植 物的抗旱性研究多在实验室和温室条件下进行,但 大田生态环境复杂,因此需进一步在自然干旱条件 下对抗氧化酶转基因植株进行抗旱性鉴定和研究; (4)植物的抗旱性最终表现在产量上,还需要对抗 氧化酶转基因植物的农艺性状指标进行鉴定。
©️ 2025 dominobio 鄂ICP备2020022707号-1  鄂公网安备42018502004957
鄂公网安备42018502004957
